3. Ciclo Celular
3. Ciclo Celular
La célula puede encontrarse en dos estados claramente diferenciados: el estado de división o mitosis y el estado de no división o interfase. Cuando la célula se divide (8) entra en el llamado ciclo celular, proceso ordenado y repetitivo en el tiempo en el que la célula proliferante crece y se divide en dos células hijas. Puede considerarse como una sucesión continua de estados que se diferencian entre sí en la cantidad de material genético existente en el núcleo de la célula (1).
Los acontecimientos más importantes dentro del ciclo celular son aquellos que tienen que ver con la replicación del genoma y la segregación de los genomas replicados en las dos células hijas formadas en la división, las cuales reciben idéntica información cromosómica (2). Estos mecanismos fundamentales se han conservado invariables a través de la evolución y ocurren de la misma forma en todos los organismos eucarióticos (9).
El ciclo celular consta de cuatro fases bien diferenciadas (Fig. 2): fase G1, fase S, fase G2 y fase M, y su duración varía entre los diferentes tipos de células eucariotas. Después de la división, las células vuelven a la fase G1, completando así el ciclo celular. Una vez en G1, las células pueden salir de este estadío y entrar en otro de descanso (G0) o bien proseguir el ciclo celular. Se puede afirmar que la formación de los dos genomas durante el ciclo celular ocurre a nivel molecular durante la fase S y a nivel celular durante la mitosis. Para asegurar el correcto desarrollo del ciclo (10), es decir, que cada nueva célula hija reciba un genoma completo, tanto el inicio y progresión de la fase S como la mitosis se controlan exhaustivamente.
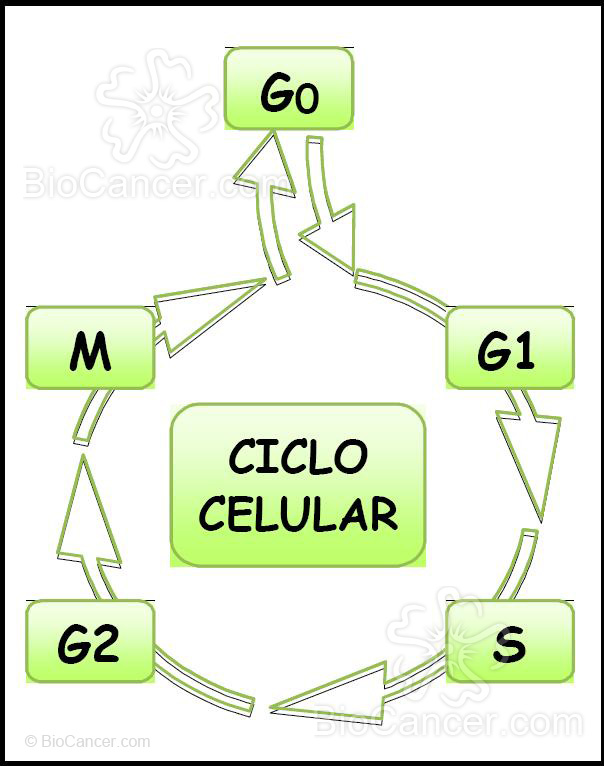
Figura 2. El ciclo celular consta de cuatro fases bien diferenciadas: fase G1, fase S, fase G2 y fase M, además de una fase G0 donde la célula no se divide. Su duración varía entre los diferentes tipos de células eucariotas.
A continuación, se describen brevemente las diferentes fases del ciclo celular (Fig. 3a-b):
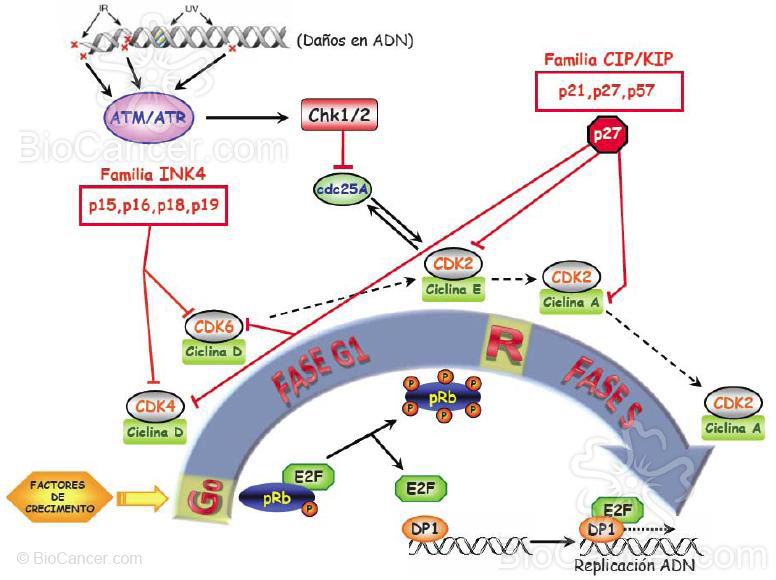
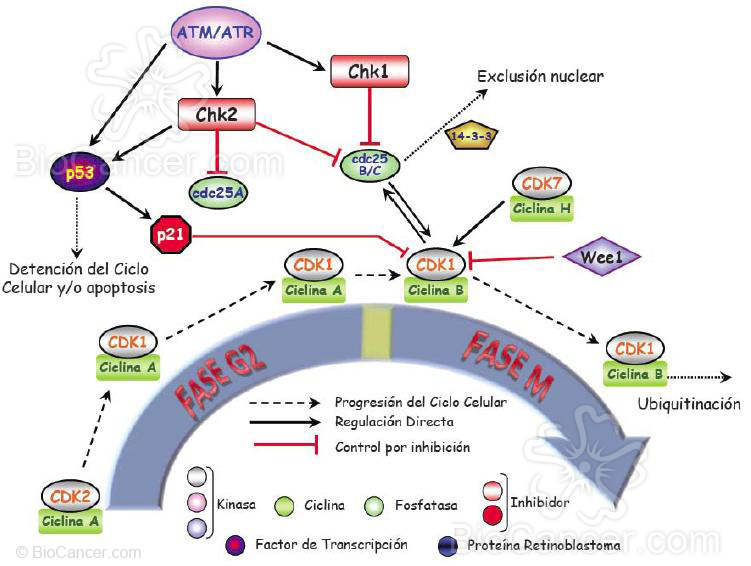
Figuras 3a y 3b. La entrada de la célula en el ciclo celular se produce cuando ésta recibe señales mitóticas como son los factores de crecimiento entre otras que la hace pasar de G0 a G1. En ese momento se activa la expresión de ciclina D que se une a las CDKs (quinasas dependientes de ciclina) 4 y 6 y de ciclina E que se une a CDK2, activándolas y produciendo la hiperfosforilación de la proteína de retinoblastoma pRb. Ésta a su vez, libera al factor de transcripción E2F y provoca la transición de la fase G1 a la S. En este punto se haya lo que se conoce como punto de restricción (R) que es el punto a partir del cual la división celular progresa independientemente de las señales mitóticas o antimitóticas que la célula reciba. En la fase S las ciclinas D y E se degradan por ubiquitinación y comienza la expresión de ciclina A que se une a CDK2. En esta fase S, el factor E2F libre se une a otro factor de transcripción, DP1, activando la maquinaria trascripcional y empezando así la replicación del ADN hasta lograr la duplicación del material genético en esta fase. Cuando esto ocurre el complejo CDK2/ciclina A inactiva a E2F y la replicación se detiene. A medida que la fase S acaba y se pasa a G2, la célula se prepara para entran en mitosis y eleva la expresión de ciclina A y B. La ciclina A se une en principio a CDK1 que se encuentra en el citoplasma pero cuando los niveles de ciclina B son lo suficientemente elevados, desplaza a la ciclina A en su unión a CDK1 y el complejo CDK1/ciclina B entra en el núcleo, teniendo lugar el comienzo de la fase M. La CDK1 se haya regulada por un lado por la fosfatasa cdc25 y por el otro por la quinasa Wee1 y por el complejo CDK7/ciclina H los cuales fosforilan o desfosforilan a CDK1 controlando así la formación del complejo con la ciclina correspondiente. Durante esta fase se produce la mitosis propiamente dicha con la separación de los cromosomas en las dos células hijas. Para que esto tenga lugar es fundamental la degradación por ubiquitinación de la ciclina B sin la cual el ciclo no continúa. Tanto la expresión de las ciclinas como la formación y función de los complejos CDK/ciclina se hallan regulados por dos grandes familias de proteínas que son capaces de inhibir ya sea a las CDKs propiamente dichas (familia INK4) o a los complejos CDK/ciclina (familia CIP/KIP). Además de estos inhibidores, en el ciclo celular existen puntos de control o checkpoints que se activan cuando se detecta daño en el ADN (ATM/ATR) que a su vez activan las quinasas efectoras (CHK1 y CHK2) y detienen el ciclo celular hasta que el daño es reparado. Para ello, activan a su vez a p53, que es la encargada de poner en marcha las rutas de reparación y, si ésta no es posible, la apoptosis.